

Back to top
Analogues d'ADN/ARN (pour l'ADN)
| Analogues ADN/ARN | Position | ||
| 5’ | 3’ | int | |
| 2’ OMe-A, 2’ OMe-C, 2’ OMe-G, 2’ OMe-U | X | ||
| 2’ MOE-A, 2’ MOE-5-MeC, 2’ MOE-G, 2’ MOE-T | X | ||
| 2’ F-dA, 2’ F-dC, 2’ F-dG, 2’ F-dU | X | ||
| LNA-A, LNA-C (5mC), LNA-G, LNA-T | X | ||
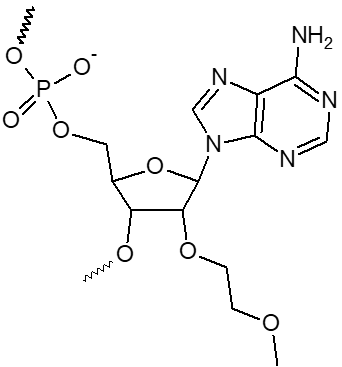
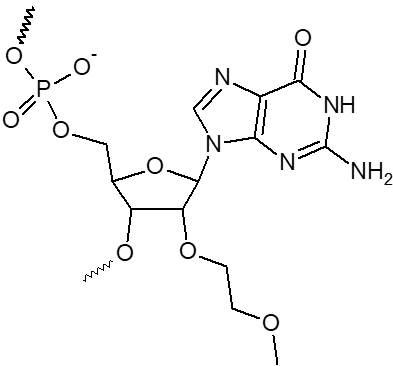
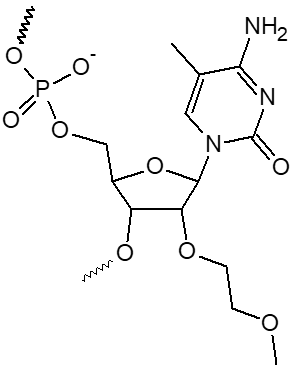
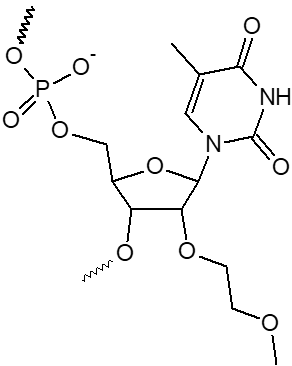
2’ OMe-A, 2’ OMe-C, 2’ OMe-G, 2’ OMe-U
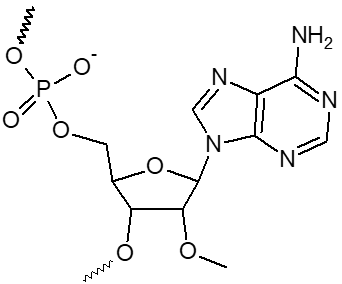
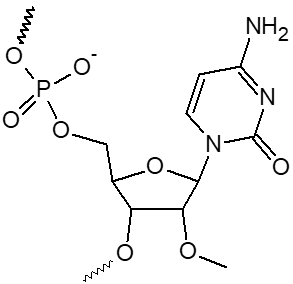
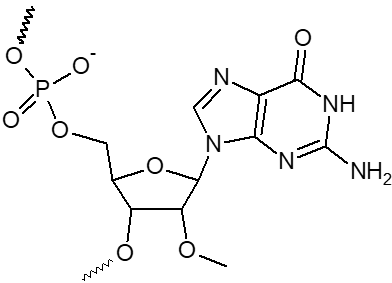
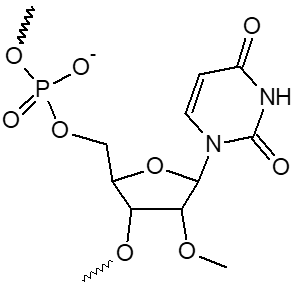
L'incorporation de nucléotides d'ARN 2'-O-méthyle procure une résistance à une grande variété de nucléases, en particulier la RNase. En outre, les oligonucléotides 2'-OMe présentent une affinité légèrement accrue envers leur séquence cible d'ARNm complémentaire, formant ainsi des duplex hybrides plus stables par rapport à leurs homologues d'ADN ou d'ARN non modifiés. Cela permet la formation d'hybrides plus stables avec des brins d'ARN complémentaires que ce ne serait le cas pour des séquences d'ADN et d'ARN non modifiées.
|
Position
|
Echelle de Synthèse [µmol]
|
Purification
|
|||||
|
|
0.04
|
0.2
|
1.0
|
15
|
Des
|
HPLC
|
PAGE
|
| Int | X |
X
|
X
|
X | X |
X
|
X |
2’ MOE-A, 2’ MOE-5-MeC, 2’ MOE-G, 2’ MOE-T
Les oligonucléotides incorporant des nucléotides modifiés par le 2'-O-méthoxyéthyle (MOE), peuvent soutenir la plupart, sinon la totalité des mécanismes d'action antisens. D'autres caractéristiques distinctives clés sont la résistance aux nucléases, une toxicité plus faible, une spécificité de liaison à la cible supérieure, ainsi qu'une affinité accrue envers l'ARN complémentaire. Pour des informations plus détaillées sur les oligonucléotides antisens 2'-MOE de Microsynth, veuillez consulter le dépliant sous la rubrique "Related Downloads".
|
Position
|
Echelle de Synthèse [µmol]
|
Purification
|
|||||
|
|
0.04
|
0.2
|
1.0
|
15
|
Des
|
HPLC
|
PAGE
|
| Int | X |
X
|
X
|
O | X |
X
|
X |
2’ F-dA, 2’ F-dC, 2’ F-dG, 2’ F-dU
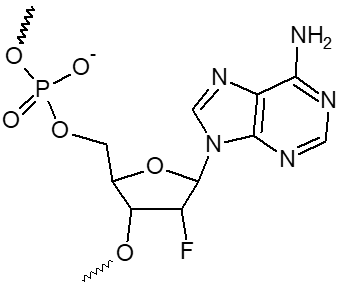
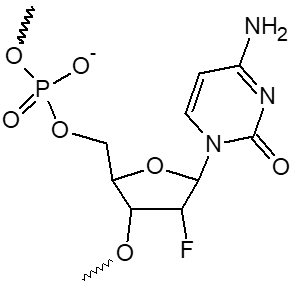
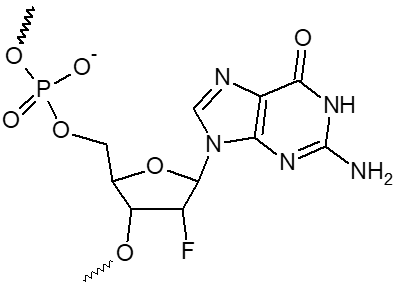
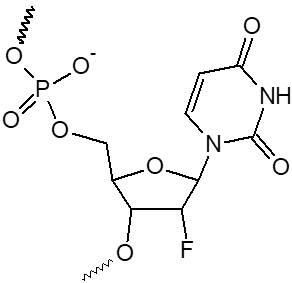
|
Position
|
Echelle de Synthèse [µmol]
|
Purification
|
|||||
|
|
0.04
|
0.2
|
1.0
|
15
|
Des
|
HPLC
|
PAGE
|
| Int | X |
X
|
X
|
X | X |
X
|
X |
LNA-A, LNA-C (5mC), LNA-G, LNA-T




|
Position
|
Echelle de Synthèse [µmol]
|
Purification
|
|||||
|
|
0.04
|
0.2
|
1.0
|
15
|
Des
|
HPLC
|
PAGE
|
| Int | X |
X
|
X
|
X |
X
|
X | |